

近年來頻傳臺灣獼猴造成各類農產品損害、傷害遊客、搶奪路人食物等人猴衝突事件,有關當局規劃各類防猴策略,如架設防猴電網、結紮、避孕、捕捉移除等控制族群數量方式,以減少獼猴族群數量降低人猴衝突事件發生。107年6月亦在專家學者考量臺灣獼猴族群量穩定及族群成長趨勢等因素下,將獼猴從保育類野生動物降為一般類野生動物。目前看來在臺灣島上兩種靈長類動物的衝突下,明明先來的臺灣獼猴卻節節敗退,在蛻下保育類物種這個保護光環後,究竟是宣判一個物種的死刑還是宣告我們對生態保育意識的成熟?
臺灣獼猴的來源
依據化石及分子證據顯示,獼猴屬生物從北非歷經數次族群擴張與遷徙事件,才抵達臺灣。臺灣獼猴最早由19世紀英國駐臺自然學家史溫侯(Robert Swinhoe)所命名,文中詳細描述臺灣獼猴的外型與習性,並與鄰近區域的獼猴比較後認為,臺灣獼猴應為獨立的物種,將其取名為
Macaca cyclopis意即圓臉的猴子(Round-faced macaca)。

史溫侯先生所在的地點就位於現今高雄壽山山腳下的打狗英國領事館,1862年的紀錄中說明,他觀察到的獼猴群偏好棲息在臨海的岩石與斜坡,以洞穴為家,對人的接近會感到十分不安並會迅速躲進洞穴中,待入侵者離去後才會再出來。此外,由十七世紀荷蘭據臺時所繪製的古地圖中,壽山地區「猴山」(Apen Berg)的地名標示,也可想見早在400多年前壽山地區的獼猴族群盛況。
在國家公園歷年的調查紀錄中顯示,在陽明山、玉山、雪霸、太魯閣、墾丁國家公園及壽山國家自然公園均有臺灣獼猴的分布紀錄,各地獼猴習性略有不同,有些會遠離人類活動區域,有些卻偏好在人類常出現的地方活動。
由史溫侯先生當時紀錄的那群在壽山活動怕人的猴子,如今卻成為時常引發人猴衝突的問題猴群,是什麼樣的原因改變了他們原本的習性?
獼猴改變習性的原因:人類餵食與人為開發
人為開發
除不當餵食行為外,開發行為也同樣劇烈的影響臺灣獼猴的族群,私自開墾的非法休息站、花園、果園及農地等,造成獼猴棲息地的分割與破碎化,原本充足的自然資源減少後,讓他們不得不與人爭地、爭奪食物資源,引發人猴衝突,造成負面形象。
在臺灣獼猴仍屬保育類物種期間,即有因負面觀感及農業損失而毒猴、虐猴、獵捕等傷害行為事件的發生,而如今獼猴成為一般類野生動物,令人擔憂傷害野生動物事件是否會更加頻繁,有賴長期監測與通報系統,來了解獼猴族群現況,確保野生族群存續。不餵食、不接觸、不干擾讓臺灣獼猴與我們能在臺灣這塊土地上共存共榮。
內政部國家公園署 版權所有 2017© All Right Reserved.
地址:10556台北市松山區八德路2段342號
")
位於在臺灣本島西南方約240海浬的珊瑚礁島─東沙島,是一個孤立的海上小島,距離最近的地區也至少相距140海浬(約260公里)。然而,根據過去的調查資料,東沙島上至少存在150種以上的維管束植物,甚至擁有一些臺灣本島所沒有或稀少的獨特植物資源,這些植物如何抵達這遺世獨立的海上孤島?又面臨什麼樣的生存挑戰?讓我們一窺其中的生命奧秘!
遠道而來的植物
漫步在東沙島海岸邊,可以看見各種姿態各異的獨特植物,從沿海慢慢地走向內陸地區,周圍植披的景觀也逐漸地從低矮的草地轉變為低矮灌叢、海岸森林的樣貌。然而在數萬年以前,腳下所踏的這片土地甫冒出水面之時,應是一片荒蕪的景象,因此科學家推測,如今在東沙島生長的植物是由島外而來。
東沙島為一沙洲島,對植物生長而言,環境並不優越。東沙島表土主要由珊瑚砂及貝殼砂組成,欠缺土壤層的結構,成為植物拓植與建立族群的重要限制因子之一,此外,東沙島冬末至初春極端乾旱缺水,亦是一個生存的嚴酷考驗。
因此如今所見的這些植物,除了要能夠順利抵達島嶼,也須克服島嶼的嚴苛物候條件,才能成功地繁衍後代,形成如今蓊鬱的樣貌。那麼,東沙島的島嶼植物是如何由島外千里迢迢而至呢?
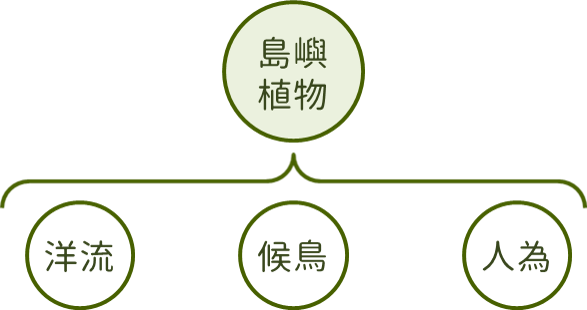
島嶼植物相主要受到自然及人為兩種方式形塑,植物可能透過季風、颱風、洋流及候鳥攜帶等自然方式抵達島嶼之上,也可能因為運送物資過程中無意間夾帶或人為刻意植栽的方式到達島嶼。
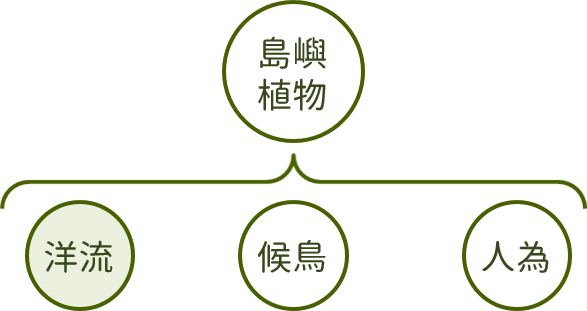
根據過去的研究成果顯示,東沙島上原生的植物有很大一部分來自「海漂種實」,這些種子和果實多具有特殊的構造得以漂浮於海面上,如種子或果皮有孔隙、子葉密度小、具有纖維或木栓層外層、結構輕盈或同時具備多種構造等,除了能漂浮在海面上外,可耐受的漂浮時間也是重要的條件。
著名的博物學家達爾文,即在1855年做過將種子浸泡於海水中的實驗,證實部分種子可耐受海水浸泡至少45天以上,仍能在播種後成功發芽,近年的實驗甚至發現部分種類甚至可耐受超過30年以上。這些種子隨著海流散布至各地的沿海區域,由東沙島周邊的洋流流向可知,東沙島的植披應與鄰近的南海島嶼及台灣西南沿海多有相似之處。
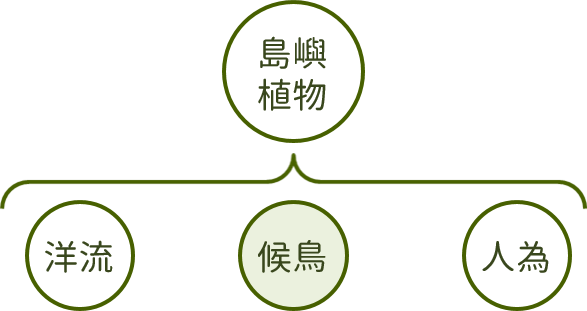
此外,候鳥的傳播也是植物播遷的路徑之一,在東沙島上倚靠候鳥傳播的植物,有些透過鮮豔多汁的果實吸引鳥類取食,再透過鳥糞傳播至島上的小葉桑、止宮樹、苦林盤等,有些則透過分泌具黏性的腺體附著於鳥羽上傳播,如白避霜花。
")
|
根據1866年英國探險船登上東沙島所留下的生態記錄顯示,當時東沙島尚處於未開發狀態,海鳥甚多,探險隊曾觀察到數以百計的白腹鰹鳥在島上礁岩的築巢育幼,但在漁民活動、20世紀初日本帝國主義擴張時期,日人侵占開採自然資源(磷礦、木材、鳥類資源等)建立南侵據點、興建各類建物後,如今此種風景已不復存在。
|
|
白腹鰹鳥
|
有鑑於此,東沙環礁國家公園亦有委託專業團隊進行「東沙島環境整理及原生植栽復育規劃」希望能逐步營造東沙島原生植披景觀。
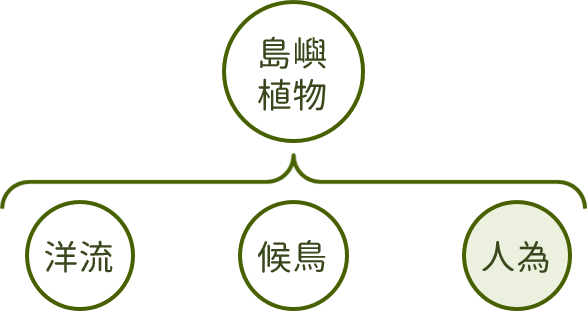
人為引入的外來物種亦不容忽視,由於島嶼相對於大陸地區而言,通常生態環境較為脆弱,較易發生物種滅絕事件,且往往難以回復原本的生態樣貌,根據過去的研究成果,東沙島上外來的植物大致可細分為:栽培種、偶遇外來種、歸化種及入侵種四類。
目前東沙環礁國家公園配合島上駐守的海巡署人員、研究人員,透過滅除、控制、減緩衝擊及持續監測等方式努力不懈地降低外來物種對原生植群的衝擊影響,並透過長期植物物候調查關注島上的特稀植物,守護這南海上美麗的指環─東沙島。
內政部國家公園署 版權所有 2017© All Right Reserved.
地址:10556台北市松山區八德路2段342號

外來物種(foreign species)
是經由人為有意或無意引入非自然分布區域的物種。外來種在移入後可能很快絕跡、成功適應新環境甚或建立穩定族群,在不受人為協助下即可在非原生地繁殖、擴散甚至造成危害的外來種則被稱之為入侵物種(Invasive species)。

臺灣外來種現況
依據
臺灣物種名錄(Catalogue of Taiwan, TaiCoL)記錄:
目前名錄中登錄有1,474種外來物種,
其中具入侵性物種有123種。
國家公園亦屬人類活動區域,難以避免存在外來物種,於本地圖上可以發現,在各國家公園區域內均有外來物種分布紀錄。
國家公園內外來種
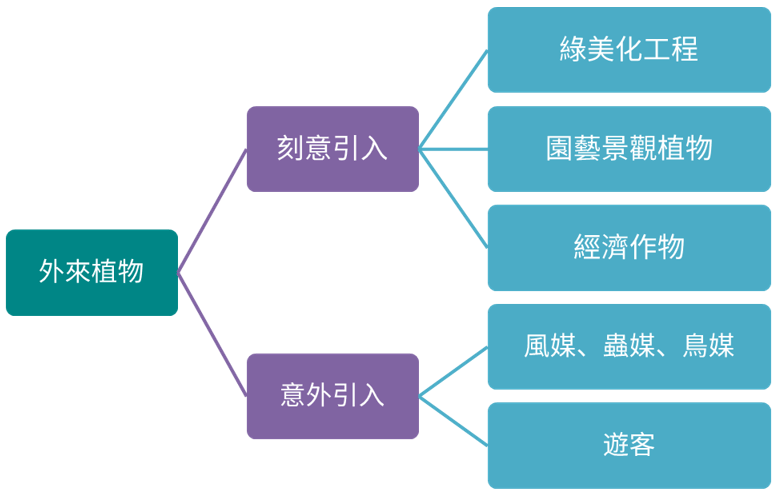
外來植物的出現與綠美化工程選用植栽、景觀植物、經濟作物、自鄰近區域藉由風媒、蟲媒、鳥媒等方式入侵…等因素有關。
國家公園內外來植物分布
因上述因素引入的外來植物,其中不乏具入侵性植物,如大花咸豐草、布袋蓮、小花蔓澤蘭、銀合歡、銀膠菊、馬櫻丹等,相關分布資料可點選各物種分布地圖,由於多數植物資源調查資料在各國家公園成立初期完成調查後較少更新,且早期資料多以名錄方式呈現,故此處之分布圖與實際危害情況存在一定程度之落差,仍有待新調查資料的補充與修訂。
國家公園內的外來動物
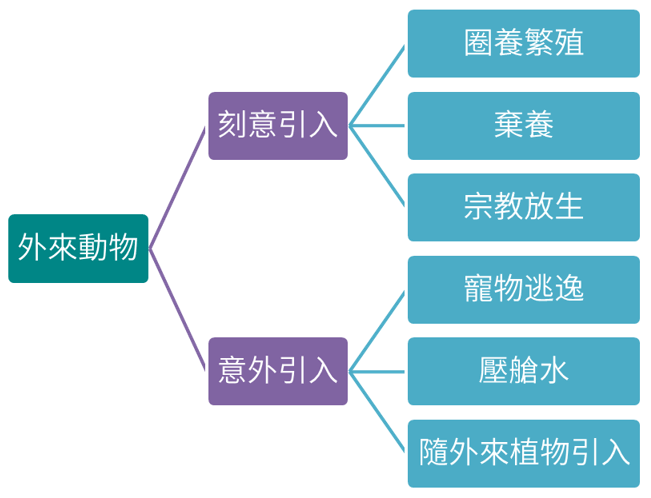
外來動物的出現與棄養或逃逸寵物、宗教放生、隨外來植物夾帶引入、壓艙水導入…等各類人類活動有關,這些因人為活動引入之外來種,可能對國家公園內原生物種產生各類不良影響,如致使當地原生物種遭受額外掠食壓力、競爭取代原生物種生態棲位、帶來疾病、造成基因汙染等各類問題。
國家公園內外來動物分布
因上述因素引入的外來動物,其中不乏具入侵性物種,如狗、貓、非洲大蝸牛、白尾八哥、埃及聖鹮等,相關分布資料可點選各物種分布地圖,此處之分布圖與實際危害情況存在一定程度之落差,仍有待新調查資料的補充與修訂。
防治外來物種入侵需要您的協助
各國家公園除委託相關專業團隊協助調查與規劃評估外來物種處置方式外,仍須仰賴民眾參與監測調查行動,目前多個管處有提供相關手冊、摺頁、解說資源等服務,歡迎您的到訪參觀。
相關監測研究計畫:
- • 壽山國家自然公園原生植物及外來入侵植物監測及分布調查計畫
- • 104年度外來種動物(兩棲、爬蟲類)入侵性調查及經營管理策略擬定
- • 壽山國家自然公園外來種鳥類長期監測調查計畫
- • 106年度墾丁國家公園龍鑾潭重要濕地(國家級)外來種魚類移除計畫
- • 107年度墾丁國家公園龍鑾潭重要濕地(國家級)外來種魚類移除計畫
- • 107年度「陽明山國家公園園區內外來種植物調查」委託辦理案
- • 墾丁國家公園區內外來植物種類及其入侵性調查
- • 106年度墾丁國家公園黃狂蟻入侵狀況調查與防治策略研擬
- • 墾丁國家公園入侵黃狂蟻監測與防治計畫

國家公園內生物資源的種類與時空分布概況,一直是各國家公園用以擬定保育策略的重要參考資料,過去各國家公園以委託專業團隊調查方式,取得國家公園內生物資源分布概略情形,呈現於
本網站中。
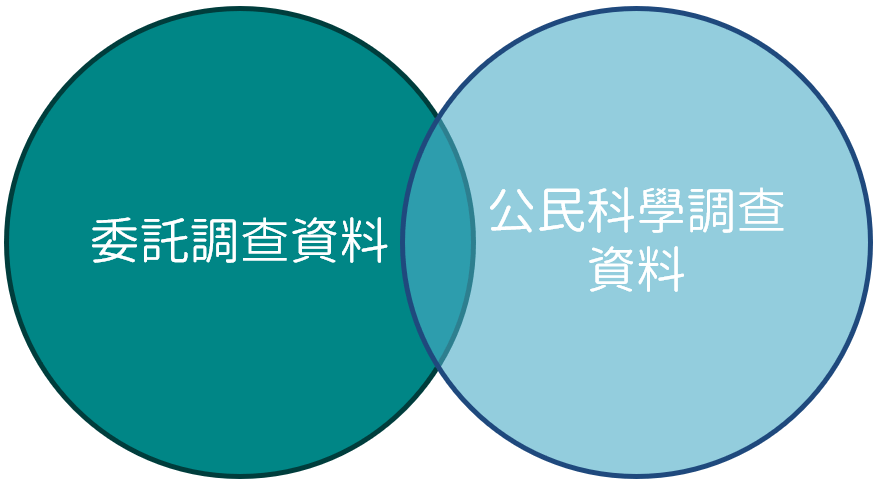
礙於須了解的資源面向廣,不同生物資源須採用不同的調查方式與委託相應之專業團隊,在經費有限情形下較難長期穩定的取得各類生物資源分布長期動態變化資料,對於氣候變遷與外來種威脅與日俱增的今日,改變現有資料蒐集模式以供擬定因應策略方式,刻不容緩,因此,國家公園生物多樣性資料庫嘗試納入公民科學調查資料,以供相關管理及研究單位作為保育策略參考依據和保育研究。
公民科學調查
公民科學調查於近年逐漸普及與精進,過去公民科學調查資料較常被詬病之處,諸如鑑定錯誤、努力量不一致、重複計數、過度估算、資料品質不一等問題,隨著大數據分析系統開發與資料蒐集模式調整下,已逐步改善。
各公民科學調查網站
近年新興崛起之公民科學調查資料庫iNaturalist便是其中一個提出資料品質控管維護的資料庫,透過使用者上傳手機或相機所拍攝之照片、影片做為佐證資料,並藉由手機或相機內建之定位系統取得經緯度座標、時間等資訊,取得穩定且可供確認之資料。

iNaturalist資料上傳機制
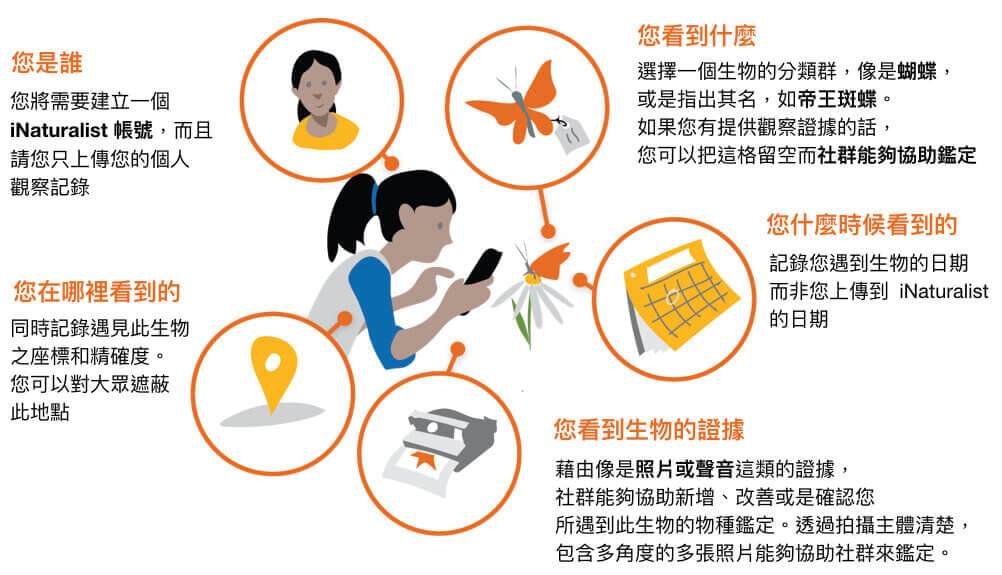
透過社群鑑定機制(Community identification)以研究等級(research grade)區分資料,確保一定程度的資料品質,十分適合用於填補現有資料的不足之處。
調查資料蒐集模式轉型
生物資源的保育若仍以早期政府主導下進行已緩不濟急,唯有逐漸轉變為全民參與模式,透過長期穩定蒐集生物資源時空分布變遷資料,才能較為正確反映現況問題,並供保育與研究單位擬定較為適切之保育策略。
台灣國家公園物種調查協作平台
本資料庫除納入iNaturalist資料外,同時開發物種調查協作平台,可供使用者查看於各國家公園內資料貢獻情形,歡迎多加利用,並共同為維護國家公園生物多樣性貢獻心力。
相關連結

環境的變遷會造成生物改變其分布範圍及族群數量。在全球氣候變遷威脅下,持續監測有多種臺灣特有生物棲息的中高海拔地區有其必要性。
高山鳥類族群變化
藉由玉山國家公園內的玉山登山步道沿線海拔3,100~3,700m的成熟林,分別於1992年、2006年及2009年調查計畫,採用相同的方法進行鳥類調查與密度估算,可作為了解高山鳥類族群變化的參考依據。
鳥類族群密度定義
對於鳥類族群密度(數量/單位面積)的變化情形,以下列方式定義:
- 鳥類族群密度「持續增加」定為「增加」
- 「持續減少」或「族群密度於2006與2009年均少於1992年的50%」定為「減少」
- 不符合上述規則者定為「波動」
玉山各鳥種的族群密度變化情形
族群密度減少
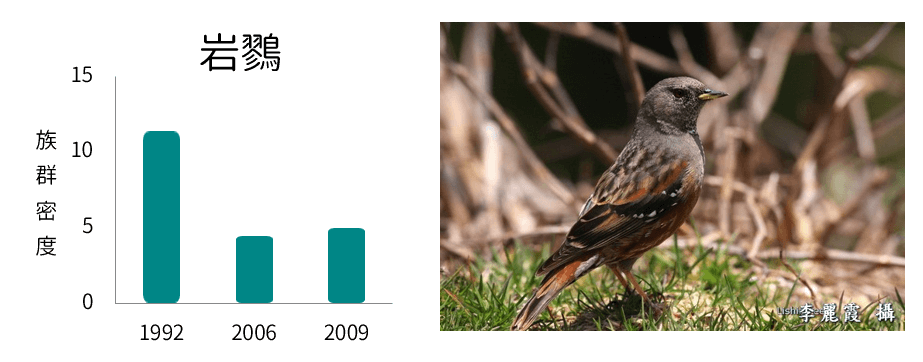
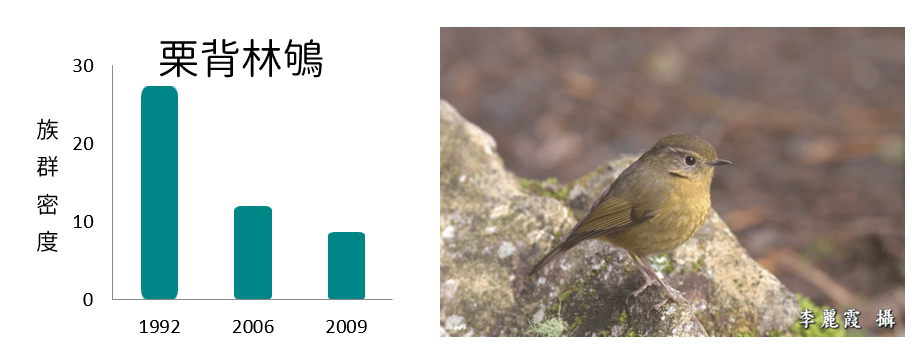
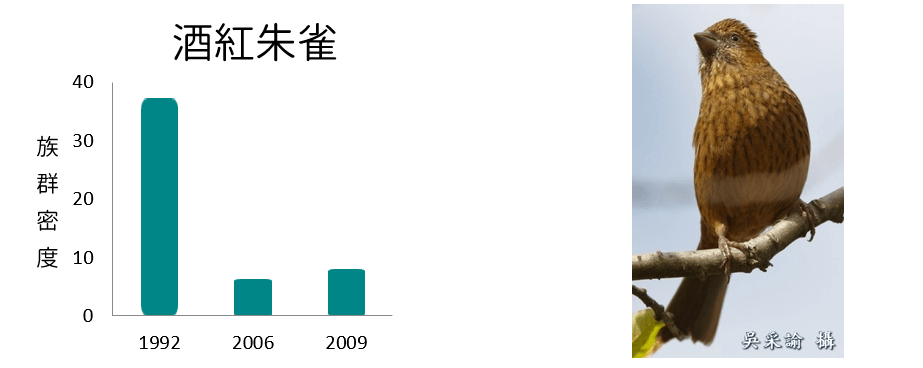


族群密度波動




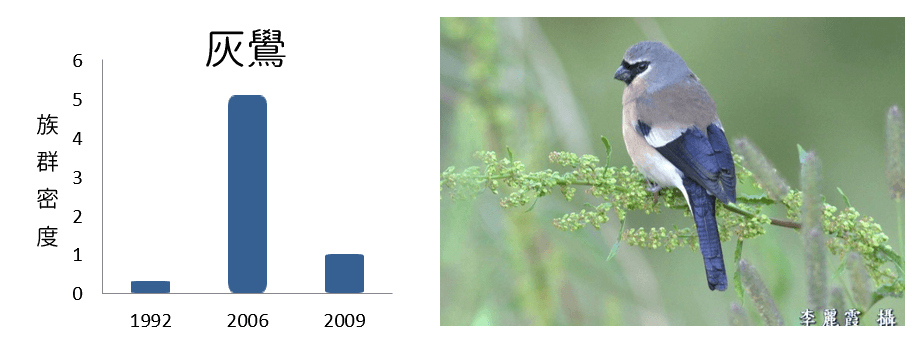
族群密度增加



從各鳥種在不同年份的族群密度變化呈現不同的趨勢變化,可以了解在環境變動下,各鳥種對於環境變動的反應不盡相同,其中族群密度減少的鳥類均為海拔下限大於2,000m,且海拔上限於3,600m以上的典型高海拔物種;族群密度增加的鳥類則為海拔下限低於2,100m,海拔上限低於3,500m的相對中海拔物種;而族群密度波動者的海拔分布則大致介於以上兩者之間。
此結果顯示鳥類的
族群密度變化與
海拔分布很可能存在著關連性,且
高海拔物種的族群可能面臨了負面衝擊。
鳥類分布海拔高度變化
除了從族群密度變化外,亦可以從分布海拔變化了解鳥類族群變動情況,由下圖可以觀察到,
三年均呈現海拔越高,鳥種豐富度越低的現象,但海拔3,600m以上的鳥種豐富度在2009年明顯較1992年增加2種以上。
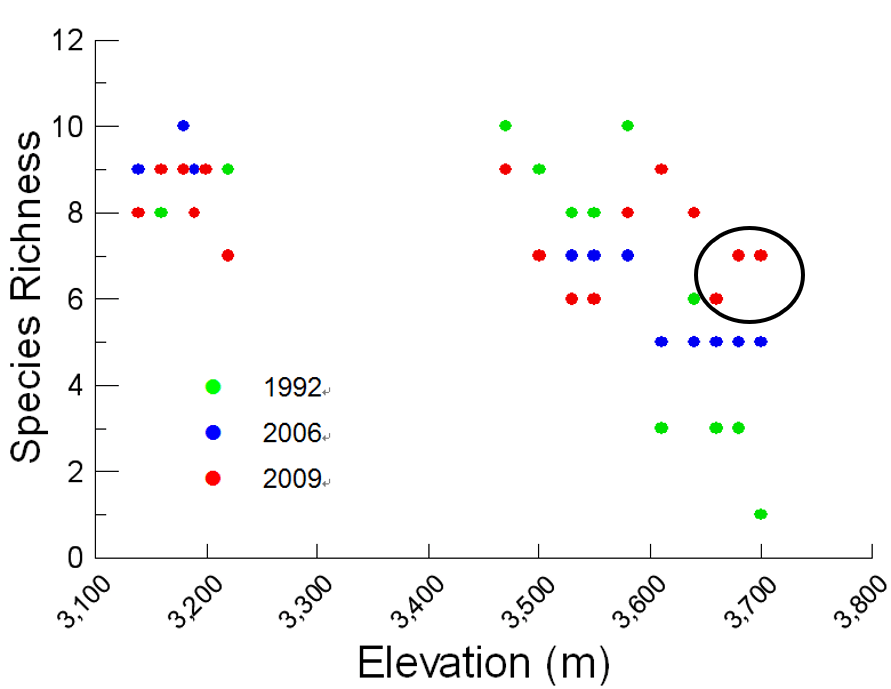
高海拔鳥類群聚變化
表、玉山高海拔鳥類群聚於各年間的平均海拔分布與上限變化情形

亦可藉由觀測平均海拔分布與平均海拔上限觀察鳥類分布是否有改變。
由於黃羽鸚嘴與茶腹鳾非三年都有出現故不列入此項分析。
餘下的11種鳥類的平均海拔分布與上限均呈現上升的情形。
表、2009年與1992年的鳥類平均海拔分布

再細部觀察此11種鳥類,海拔平均分布上升者共有9種,海拔分布不變或略降的有2種(鷦鷯與臺灣噪眉);而9種海拔分布上升的鳥類中有5種的海拔差異在平均值75m以上,包括白眉林鴝、灰頭花翼、深山鶯、火冠戴菊鳥,以及煤山雀。此外,因非三年均有資料而為納入海拔分布分析的黃羽鸚嘴與茶腹鳾均為相對海拔分布較低的物種,在1992年以前沒有在涵蓋的海拔範圍(3,100m以上)發現,而分別在2006年與2009年開始分布達3,200m,亦為鳥類海拔分布上升的現象。
結語
綜合以上,由三年的監測資料可以瞭解玉山高海拔鳥類群聚的族群與分布兩種層面的變化情形。在族群方面,顯示高海拔鳥種族群可能面臨了負面衝擊,而在分布方面,鳥類沿海拔梯度的分布情形在年間有差異,近年更有朝較高海拔移動的現象。
本範例雖然時間尺度橫跨長達17年,但實際僅有3年,亦即3個時間點的資料,考量生態系中存在的變異性,這樣的非連續觀測資料對於瞭解趨勢與證據說服力仍有很大的加強空間。這也建議著「長期監測」的重要性。
參考文獻
- • 丁宗蘇. 1993. 玉山地區成熟林之鳥類群聚生態. 碩士論文. 國立臺灣大學, 台北.
- • 許皓捷. 2003. 臺灣山區鳥類群聚的空間及季節變異. 博士論文. 國立臺灣大學, 台北.
- • 李培芬. 2009. 玉山鳥類族群監測. 空間生態實驗室調查資料. 國立臺灣大學, 台北.